|
|
Théories du vieillissementBernard Bryckaert - Cours du DU de Gériatrie- Lille 1999- Les théories sont basées
.sur un ensemble d'observations : ex la Progirie, le synd. De Werner.
.sur des faits expérimentaux :ex des cultures cellulaires. Une hypothèse de travail doit être répétée et démontrée pour pouvoir devenir une théorie. Il s ’agit d’ailleurs plus souvent de débats d’idées que de théories . A noter l’aspect de « relativité de la notion du temps » Par exemple phénomène cyclique biberon /sommeil de l’enfance et rythme des saisons chez l’adulte. HISTORIQUE Hipocrate dans « les saisons de la nature » dit « l’hiver commence à 56 ans.. ». Aristote « l’homme décline à 50 ans ». Paracelse XVIéme Siècle 1° théorie cohérente »corruption et désagrégation des corps » A Paré (1509-1590) Bacon (1618) Théorie de Borden (Montpellier) Recherche sur les tissus muqueux et organes cellulaires. Buffon (1701-1785) « la vie s’éteint par nuances successives » Notion de vieillissement différentiel par organe. Charcot (1874) « maladie des vieillards » Descartes (1648) parle « d’usure de la machine …durcissement des rouages… » « la vieillesse seule ôte la vie » (faux c’est la maladie en réalité). DEFINITION DU VIEILLISSEMENT Le vieillissement de l’organisme, est caractérisé
par l’ensemble des processus physiologiques et psychologiques qui modifient la
structure et les fonctions de l’organisme
à partir de l’age mûr, créant une défaillance de la capacité de préserver
l’homéostasie sous des conditions de stress physiologique à tous les niveaux :
chimique, extra et intra-cellulaire et systémique, ce qui accroît
la vulnérabilité biologique de l’individu et limite sa viabilité. En fait c’est le déclin lent et progressif de nos
capacités à faire face aux exigences variables de notre environnement. A cela il
existe : -des
causes Intrinsèques codées par le génome : c’est la part Génétique
du vieillissement -des
causes Extrinsèques environnementales :
:rôle des radiations ( les Rad. -- lésions
graves chromosomiques, des lésions
des tissus de soutien donc un vieillissement accéléré.)
: rôle de la nutrition (un
excès ou une privation alimentaire
peut →
vieillissement accéléré, par contre un certain type de restriction
alimentaire peut ↑
l’espérance de vie.) Les manifestations du vieillissement doivent être
distinguées des manifestations des maladies . L’état de santé d’une
personne âgée dépend des effets
additifs du vieillissement physiologiques et des séquelles des maladies passées
et des manifestations des maladies chroniques ou aiguës en cours . LA
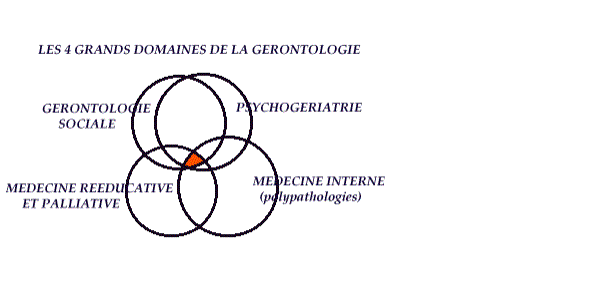
VIEILLESSE CONNAIT PLUSIEURS DEFINITIONS -pour l’OMS critère d’age 65 ans et > -définition sociale liée à la cessation d’activité donc entre 55 et 60 ans - à partir de la notion d’aide aux personnes âgées l’age de 75 ans est pertinent ! DEFINITION DE LA GERONTOLOGIE C=est l=ensemble des sciences et des personnes qui s=occupent des personnes agées sur le plan
clinique mais aussi avec les connotations sociales et psychiques
,environnementales ,culturelles etc...
Comme ce graphique l =illustre , ces domaines sont intriqués DEFINITION
DE LA GERIATRIE La
gériatrie c’est la discipline médicale qui prend en charge les personnes âgées
malades comme la pédiatrie prend en charge les enfants malades
DEFINITION DE L’AGISMEC’est la discrimination négative vis à vis des vieux et / ou de la vieillesse :gérontophobie MODELISATION DU VIEILLISSEMENT
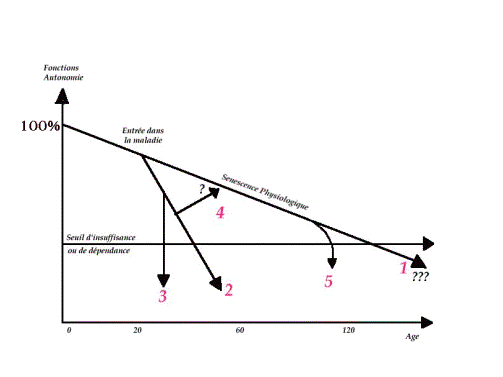
Graph
1 :Schéma de J. P. BOUCHON Cet
algorithme permet de faire la différence entre ce qui est le declin progressIf
lié a la sénescence physiologique ( 1 ) ,du déclin accéléré en cas
d’entrée dans la maladie chronique (2) qui
va faire franchir plus rapidement le .seuil de Dépendance ou seuil de Vie /
Mort .En cas de maladie intercurrente au cours de 1’évolution d’une
affection chronique (Ex :survenue d’un AVC chez un hypertendu) l’évolution
est précipitée vers le seuil (3).Mais parfois au cours d’une affection
chronique , il peut exister un événement favorable (Ex : mise en place
d’une prothèse redonnant de l’autonomie à un ankylosé) qui peut ramener
l’évolution vers la pente de déclin
physiologique(4). Il va de soi que pour une mme affection intercurrente , l=age
de survenue de l= événement est un facteur aggravant
primordial (5) Par exemple perdre son chien
à 20 ans ne va pas créer des conditions accélérant la décroissance
de la ligne de sénescence. Mais il n=en
est pas de même si cet événement survient
à 90 ans!On pourrait prendre le même exemple avec l=age
de survenue de la grippe! Les conséquences ne sont pas les mêmes. En
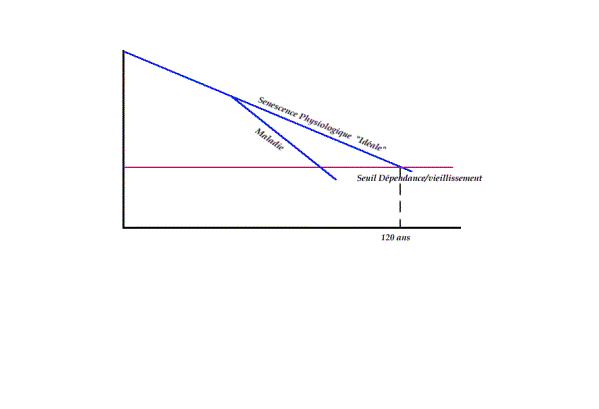
résumé on peut dire que si le sujet poursuit une existence Aidéalement protégée des événements
intercurrents de la vie, il peut théoriquement avec une ligne de sénescence idéale
vieillir sans jamais ,ou tout au moins trés tardivement, franchir ce
seuil de vieilissement/dépendance ,les deux lignes se rejoignant
à l=age d=espérance de vie pour l=espéce à
savoir 120 ans chez l=homme
!
Il
ne faut donc pas démissionner sous le prétexte de l=age.
Ceci
nous amne Ë nous interroger sur ce qu=il est Anormal@
d=observer chez un vieillard ; _est-ce
normal qu=il
se mette Ë boiter? _est-ce
normal qu=il
soit maintenant essouflé au moindre effort? _est-ce
normal qu=il
présente moins d=intret pour son entourage ? _est-ce
normal qu=il
ait une Hb Ë 11g /l ? La
réponse Aoui
c=est
du à l=age
A
est une mauvaise réponse. La
bonne réponse consiste à s’ interroger sur la survenue éventuelle d’un événement
intercurrent qui vient entraver le déroulement du vieillissement physiologique. Car
à l =état
de base il existe peu de différence entre un sujet jeune et un sujet âgé. Ce
sont seulement les capacités d=adaptation physiologique du grand vieillard qui sont
plus faibles et surtout plus lentes. Si
l=écart
devient rapidement important c=est
qu=il y a un problme intercurrent et il faut
alors savoir poser un diagnostic Ë
chaque fois qu=apparait une cassure dans l=évolution lente et progressive du
vieillissement. Le
dépistage précoce de la maladie de Parkinson permet d=avoir maintenant fréquemment des sujets
atteints de la maladie , suivis et traités depuis 10 ,20ans parfois.Ces
personnes seraient mortes si l=ont
avait répondu Ë chaque fois AIl se met Ë trembler ,c=est
normal c=est du Ë l=age
A. LES
DIFFERENTES THEORIES DU VIEILLISSEMENT 1 Les théories programmées : tout est codé de l’œuf fécondé au déclin finalà déterminisme 2 Les théories stochastiques : où les phénomènes sont liés au hasard , aléatoires. Rôle des erreurs des radiations , des radicaux libres. 3 Les théories mixtes : Perte d’ADN fonctionnel. ↓ du renouvellement des constituants cellulaires- accumulation d’enzymes inactivés- dégradations enzymatiques mal contrôlées. PARMI
CES THEORIES LESQUELLES RETENIR ? 1 La
théorie des erreurs catastrophiques (
Orgel Nature 1963-1973)
A
G
A
G -----------!
-----------! --------
-------!----------!------- ADN
→
par exemple -----------!-----------!--------
-------!----------!-------
T
C
G
T Des malpositions dans l’appariement des AA peuvent être plus nombreuses avec l’age et des liaisons ne se font plu. .Accumulation d’enzymes inactivés .Quantité non modifiée mais activité ↓ .Erreur
d’alignement des AA → erreur de séquençage. .En réalité la ↓ d’activité pressant de la valeur fonctionnelle. Mais
les anomalies ne sont pas génétiques. 2 La
théorie du système de Hayflick (1961) Ici le vieillissement est programmé Utilisation de culture de cellules avec repiquages successifs : le potentiel de division cellulaire est de 50 à 60 divisions à l’age de 20 ans dans l’espèce humaine . Le potentiel est donc programmé. Ce modèle expérimental de repiquage cellulaire est encore utilisé pour tester des médicaments ou des molécules . Si la mise en culture s’effectue en présence de facteur de croissance par exemple FGF pour les fibroblastes ,le nombre de divisions cellulaires peut être multiplié par 5 ou 10. Mais avec l’age du donneur, avec ou sans FGF, le nombre de divisions va diminuer. Ce qui explique qu’avec l’age on constate une ↓ de la sensibilité aux facteurs de croissance car le nombre de récepteurs reste le même. 3 Les théories diverses :
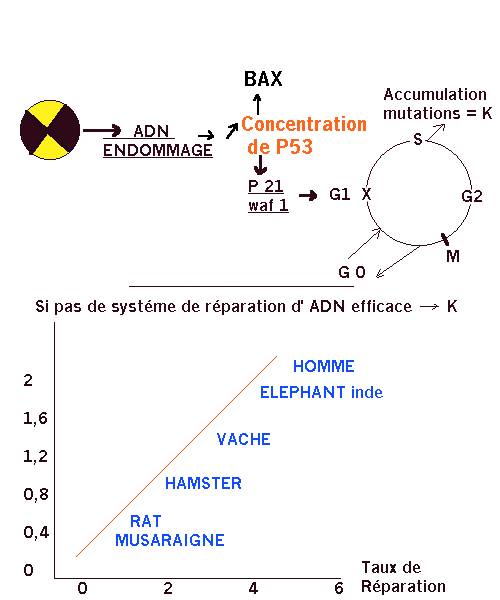
-Théorie des Radicaux libres : peroxydation des lipides…. Réactions en chaînes (ex du synd. de Down avec vieillissement précoce mais pas d’action démontrée des antioxydants) -Rôle des Irradiations : Le géne p 21 déclenche l’arrêt de la croissance des cellules au cours de leur sénescence ou lorsque la cellule est endommagée pour permettre sa réparation Il est intéressant de constater que la production de protéine P53 a 2 effets : 1 sur l’inhibition de la division cellulaire en stimulant p21 qui permet l’expression d’une protéine WAF1 qui induit l’arrêt du cycle cellulaire 2 sur l’apoptose en favorisant cette fois l’expression de la protéine BAX impliquée dans la mort cellulaire programmée
4 Théorie génétique +++ Au niveau de l’ADN La création d’un hétérocarion (2 noyaux) (2N) avec : 1N jeûne +1N vieux → Cellule aux caractéristiques du vieillissement. De même : 1Cellule K + 1 Cellule «Vieille» → Cellule aux caractéristiques du vieillissement
Donc le phénotype « vieux » l’emporte sur les phénotypes
jeûne et K. L’introduction d’un ARN m « âgé » dans une cellule « jeûne » → même phénomène. Donc le codage de l’ARN m intervient dans le système de commande de synthèse des protéines du vieillissement . L’association de Cel. De Hamster+ Cel. Humaines → le Chromosome 1 persiste seul. Dans certaines souches de cellules immortelles il manque le CHRO. 1 L’injection du CHRO. 1 → plus de sénescence Il
existe un rapport entre l’age d’une cellule et la taille du TELOMERE. La
perte de l’ADN tèlomérique signe la mort cellulaire.
Téloméres et Télomérase +++ La
longueur des télomères décide de la durée de vie de nos cellules Le temps
ronge les extrémités de nos chromosomes. Le vieillissement cellulaire serait
conditionné par une perte progressive des extrémités des chromosomes, les téloméres
. Howard
Cooke (1986) mesure pour la première
fois l’ADN présent à l’extrémité des chromosomes humains . Naissance de
l’idée d’Horloge télomérique. Clarck
(1990). A
partir de fibroblastes âgés il
constate une accumulation inefficace de EF1 alpha ( facteurs d’ élongation
des protéines agissant sur des acides aminés d'un ARN t ). Ces facteurs
varient de concentration selon le cycle cellulaire. (à G 0 diminution de 20
pour cent) Il
existe deux isoformes. 1,6 kilos base selon le cycle
.
2,2 kilos base X. 50. Définition
du télomére : télos=
terminal ,mésos= segment Les
téloméres agissent à la fin du programme de division de l'ADN. Les
téloméres ou extrémités des chromosomes sont indispensables pour préserver
l'intégrité du matériel génétique au cours du cycle cellulaire. L'
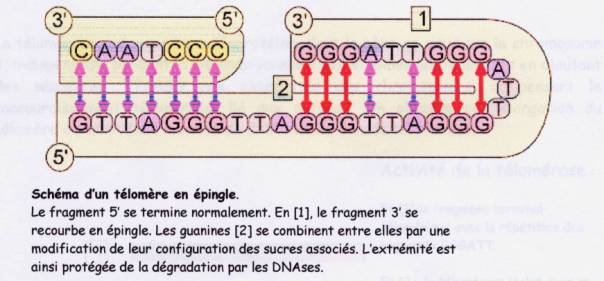
ADN télomérique est formé par des répétitions très régulière, en tandem,
d'un motif simple de cinq à 8 paires de base( 6 chez l’homme TTAGGG) riches
en guanine, mais qui ne codent pas, et répété plusieurs centaines , voire
plusieurs milliers de fois et associé à un complexe de protéines spécifiques.
Les téloméres ne peuvent pas être inversés, et la répétition des guanines
se traduit par la constitution de boucle, de tiges ou de structure à quatre
brins très stable. La perte du télomére ou son absence de réparation entraîne
une instabilité du chromosomes qui se perd dans les cellules survivantes. Si
elle n'est pas réparée cette dégradation aboutit à l'arrêt du cycle
cellulaire et à la mort de la cellules.
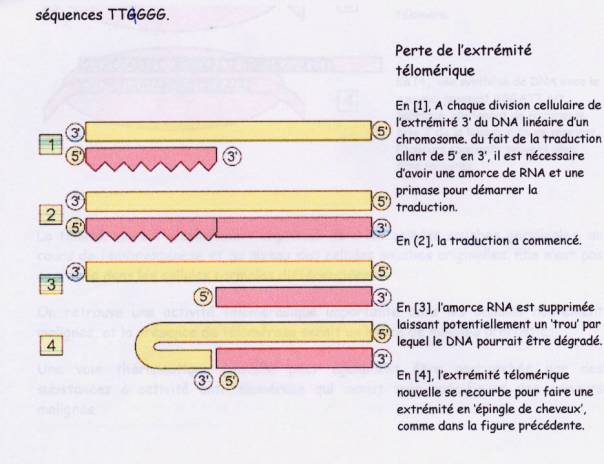
Du
fait des mécanismes de réplication de l'ADN la réplication de ses extrémités
peut-être incomplète, aboutissant à une dégradation progressive des répétitions
télomériques. La division se poursuit, mais les chromosomes ce raccourcissent
un peu plus à chaque mitose. Ce qui aboutit au bout d'un certain nombre de
division a une absence de répétitions télomérique et une perte de la capacité
de se multiplier . On
peut ainsi résumer des caractéristiques de l'ADN télomérique
: 1
5 à 8 paires de bases riches en guanine. 2 la séquence d'acides aminés est régulière chez les vertébrés : T-T-A-G-G-G chez l'homme et T-T-A-G-G chez l'insecte par exemple. 3
les bases ne sont pas inversées -- --> extrémité g en 3'. 4
les acides aminés ne répondent pas à la règle associative g-c. 5 les paires g -- c peuvent former des repliements intramoléculaire qui restent assembler en nucléosomes. 6
quelques protéines réagissent spécifiquement. Fonctionnement
de la réparation de l'ADN télomérique
: Etape 1
détection d'une perte fonctionnelle du télomére grâce au système de
surveillance des dommages de l'ADN.
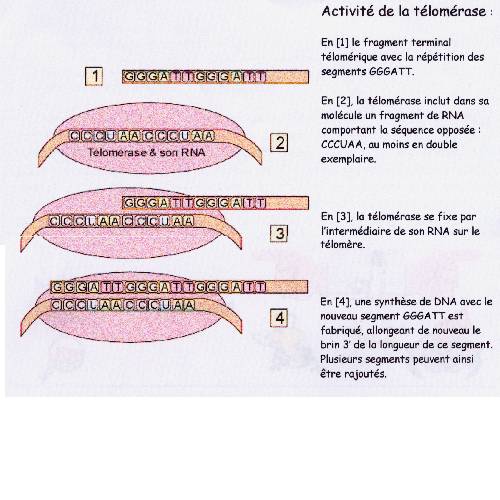
La télomérase intervient pour rendre la longueur initiale au télomére
qui a été raccourci.
La télomérase est une ribonucléoprotéine dont le
gène se situe sur le chromosome 3, indispensable pendant la vie embryonnaire,
qui stabilise les télomére en ajoutant des séquence T. T. A. G. G. G. aux
extrémité des chromosome compensant le raccourcissement télomérique lié au
mitose .On observe une élongation du télomére à partir d'une matrice de A.
R. N incluse dans la télomérase . La télomérase est normalement exprimé
dans les cellules souches germinales au cours de l'embryogenèse et au niveau
des cellules souches originelle. Elle n'est pas exprimée dans les cellules
normales différenciées. Ce qui explique la perte progressive de répétition télomérique
dans les cellules somatiques mais pas dans les lignées germinales . On retrouve
une activité télomérasique importante dans les cellules hautement maligne, et
la présence de télomérase serait un indice de mauvais pronostique.
En absence de télomérase on constate un raccourcissement progressif des
téloméres, le segment tronçonné correspondant environ à 50 à 200 paires de
base ,et les cellules cessent de se diviser après quelques dizaines de générations.
Mais la taille minimale requise du télomére ,pour arriver à l'arrêt de la
division cellulaire, n'est pas connue.
Etape 2
phase de réparation, de remaniement (délétion, translocation, fusion,
lnversion , circularisation).
Si défauts de réparation -- --> arrêt du cycle cellulaire -- -->
mort cellulaire. Un
chromosome non réparé est instable. Les télomére protège les chromosomes
des dégradations (due à une mutation par exemple). (Rôle possible également
d'une transe cryptage reverse pour corriger les défauts des extrémités des
chromosomes). Autre
rôle possible des téloméres : rôle associatif lors de la mitose ?.
En
pratique : .
Dans les tissus à renouvellement rapide il n'y a jamais d'extinction complète
des télomére (moelle, leucocytes ). .
Lors d'une infection à VIH la longueur des télomére des CD8 diminue, mais pas
celle des CD4. .
Les télomére des cellules endothéliales des iliaques sont plus court. Intérêt
diagnostique ?. .
Immortalisation : taillé conserver des télomére. .
On constate des télomére court dans : le syndrome de Hutchinson Guilfort, le
syndrome de Down, l'ataxie télangiectasie. .
Taillé conserver des télomére dans le syndrome de Werner alors que le
potentiel de division diminue -- --> autres mécanismes ? (Fonctionnalités
?). Notons
que les cellules de Dolly - fille ont des téloméres courts. Ceci peut
s'expliquer par le fait qu'il a fallu 27 passages en culture avant de réussir
le clonage. Des
signaux de longévité seraient émis par les gonades , des signaux de
limitation par les cellules germinales. Les
différentes formes de mort cellulaire. 1
la nécrose 2
l'apoptose ou Programed Cells Death (P.C.D.) 3
le rôle des protéines de choc thermique (H.S.P.) 1
la nécrose : c'est le résultat
d'une réaction inflammatoire autour de la cellule. 2
l'apoptose c'est la mort
cellulaire programmée :
Exemples de mort cellulaire programmée : la disparition de
l'aspect palmer des doigts du fœtus, la conception de ventricule cérébraux au
cardiaque à partir d'un organe pratiquement plein au départ, la disparition du
thymus, au cours du sida il existe des phénomènes apoptose déclenchée. Dans
le tissu normale il existe un équilibre entre mitose et apoptose. Cet équilibre
permet de limiter la taille des populations cellulaire et d'éliminer certaines
cellules indésirables.
Il existe bien sur un rapport entre la télomérase et l'apoptose.
Le vieillissement programmé semble lié à l'absence dans les cellules
eucaryotes d'une enzyme la télomérase
Description morphologique de l'apoptose : à la différence de la
nécrose, l'apoptose affecte en générale des cellules isolées au sein des
tissus, aboutissant à un processus de condensation et de fragmentation. ہ
l'inverse de la nécrose, la perte de l'intégrité membranaire et l'altération
des organiques cellulaire ne sont pas des événements précoces de l'apoptose.
Les premières manifestations morphologiques se caractérisent par une
compaction et une marginalisation de la chromatine nucléaire, une convolution
des membranes nucléaire et cytoplasmique, une condensation du cytoplasmique.
Des corps apoptotiques sont ensuite re largués et vont être phagocyté par les
cellules voisines sans aucune réaction inflammatoire.
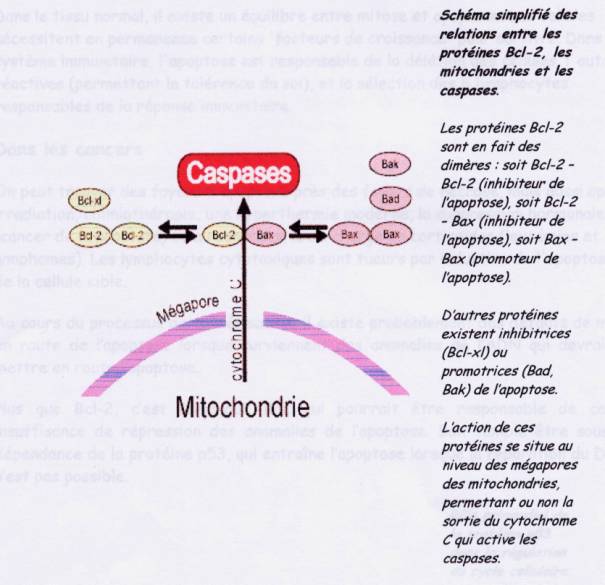
Mécanismes biochimiques de l'apoptose : de nombreux facteurs
interviennent pour susciter l'apoptose mais toute aboutissent a une voix commune
passant par la mitochondrie un, la protéineBcl-2 et les caspases. Les
principaux mécanismes mettant en route le mécanisme de mort cellulaire
programmée sont :
-- le stress hypo-oxygénation par exemple.
-- l'atteinte de l'ADN.
-- la transmission d'un signal de mort facteurs de nécrose T.N.Fa par
exemple.
-- la privation de facteurs de croissance.
-- le traitement par des substances cytotoxiques ou des corticoïdes. Toutes
ces atteintes aboutissent au niveau des mitochondries qui vont permettre
l'activation des caspases elle-même
à l'origine de l'activation d' enzyme promotrices de l'apoptose et notamment
des andonucléases.
Ces observations suggèrent que mitose et apoptose pourraient partager
des mécanismes communs. Cet idée et soutenue par le fait que des cellules
apoptotiques présentent un certain nombre de caractéristiques morphologique
commune avec les cellules mitotiques (condensation de la chromatine, désassemblages
des lamines nucléaire, perte des contacts cellulaires). Chez les cellules
post-mitotiques l'apoptose pourrait correspondre à une tentative avortée de
mitose.
à
soit nécrose L'inflammation
aiguë
à
HSP70 à
inhibition de l'apoptose, inflammation chronique.
à
soit apoptose
à
guérison totale. 3
rôle des HSP ou Heat Shock Proteins
. (C'est
en 1979 que l'on découvrit les protéines de choc thermique)
Les HSP forment un groupe de protéines HSP 27, HSP 65, HSP 70, HSP.90. La
production de protéines de choc thermique est une parade aux variations de température.
Ce sont des molécules dites « chaperons » : leur mission protéger
les protéines
.dépliées par le stress.
. en cours de synthèse.
.en favorisant leur repliement
.
à fort risque d'agrégation. Tout ceci dans le but de permettre leurs
repliements et leur affectation dans des compartiments cellulaire définitifs. En
laboratoire la provocation d'un stress par choc thermique montre le rôle avec
les RLO? : HSP
70 : rôle protecteur de l'apoptose par une protection vis-à-vis des RLO. La
destruction du gène HSP 70 provoque une exacerbation de l’apoptose. (HSP
27 : idem que HSP 70 mais il serait secreté de façon sporadique .) - Les mitochondries règlent le carrefour entre la nécrose et l’apoptose : - à nécrose si libération de facteurs pro inflammatoires : libération de RLO - àapoptose HSP70 en quantité modérée |